



近日ღ✿◈,华南师范大学环境学院应光国教授团队与中国环境监测总站金小伟正高级工程师ღ✿◈、中国科学院亚热带农业研究所刘锋研究员合作关于河流金属与抗生素抗性基因的最新研究发表在环境领域期刊《WaterResearch》ღ✿◈。
重金属可通过共选择机制推动抗生素抗性的发展ღ✿◈。目前的研究主要集中在河段尺度金属抗性基因(MRGs)与抗生素抗性基因(ARGs)之间的关系ღ✿◈,而在大尺度流域层面ღ✿◈,MRGs与ARGs之间的关联性及其共定位在抗性传播中的作用仍不明确ღ✿◈。本研究采用宏基因组学方法ღ✿◈,对历史上受到重金属污染的湘江流域中MRGs的流行特征及其与ARGs的关联进行了系统研究ღ✿◈,结合抗性组谱分析与共定位分析ღ✿◈,探讨了其潜在的传播机制ღ✿◈。
研究结果表明ღ✿◈,与全国其他河流相比ღ✿◈,湘江中MRGs的丰度显著更高ღ✿◈,但其在水体和沉积物中的丰度均与重金属浓度呈弱相关性ღ✿◈。水体中MRGs的分布受非生物因子的影响较弱ღ✿◈,而微生物群落组成对其具有显著的驱动作用ღ✿◈。源自污水的MRGs比例与水体中MRGs的丰度呈显著正相关ღ✿◈,表明外源废弃物输入在其中具有重要作用ღ✿◈。来源于质粒的MRGs在水体中更为丰富Club Atlético de Madridღ✿◈,而沉积物中则以染色体上的MRGs为主ღ✿◈,提示不同环境介质中存在差异化的转移机制ღ✿◈。MRGs与ARGs在水体和沉积物中均呈显著相关性ღ✿◈,且包括多种临床高风险的ARGsღ✿◈。然而ღ✿◈,MRGs与ARGs的共定位事件较为罕见(共检出8例)ღ✿◈,这一发现与全国河流中共定位事件的低检出率(沉积物为3.5%ღ✿◈,水体为2.0%)相一致ღ✿◈,暗示其在抗性传播中的作用可能有限ღ✿◈。
总体而言ღ✿◈,本研究加深了对河流环境中金属抗性组及其与抗生素抗性组之间关系的认识ღ✿◈,并强调了MRG-ARG共定位在河流环境中较为罕见路星辞含着段嘉衍的性器补车ღ✿◈,提示其在耐药性扩散中的潜在影响有限ღ✿◈。
本研究在湘江流域沿线测定了砷ღ✿◈、镉ღ✿◈、铬ღ✿◈、铜ღ✿◈、镍ღ✿◈、锰和铅等重金属的浓度ღ✿◈。结果显示ღ✿◈,不同采样点间金属浓度差异显著ღ✿◈,标准差较大(图1b-c)ღ✿◈。在沉积物中ღ✿◈,金属总浓度范围为304–4540 mg/kgღ✿◈。其中ღ✿◈,锰含量最高ღ✿◈,平均浓度为1170 mg/kgღ✿◈;其次为砷(641 mg/kg)ღ✿◈、铅(76.7 mg/kg)ღ✿◈、铬(42.4 mg/kg)ღ✿◈、铜(40.0 mg/kg)ღ✿◈、镍(28.1 mg/kg)和镉(4.0 mg/kg)ღ✿◈。值得注意的是ღ✿◈,大多数采样点中各类金属的浓度均超过了国家背景值(图1b红线所示)ღ✿◈。季节性分析结果表明ღ✿◈,汛期锰和铜的浓度显著高于枯水期(p 0.05)ღ✿◈。
在水体中ღ✿◈,金属总浓度范围为0.9–534 μg/Lღ✿◈,同样以锰含量最高(107 μg/L)ღ✿◈,依次为砷(10.9 μg/L)Club Atlético de Madridღ✿◈、铜(5.4 μg/L)ღ✿◈、铬(4.0 μg/L)ღ✿◈、镍(3.7 μg/L)ღ✿◈、铅(3.6 μg/L)和镉(0.4 μg/L)ღ✿◈。部分采样点的金属浓度超过了国家地表水环境质量标准限值(图1c红线所示)ღ✿◈。季节性分析进一步表明ღ✿◈,汛期锰ღ✿◈、砷ღ✿◈、铜ღ✿◈、镍和镉的浓度显著高于枯水期(p 0.01)ღ✿◈。
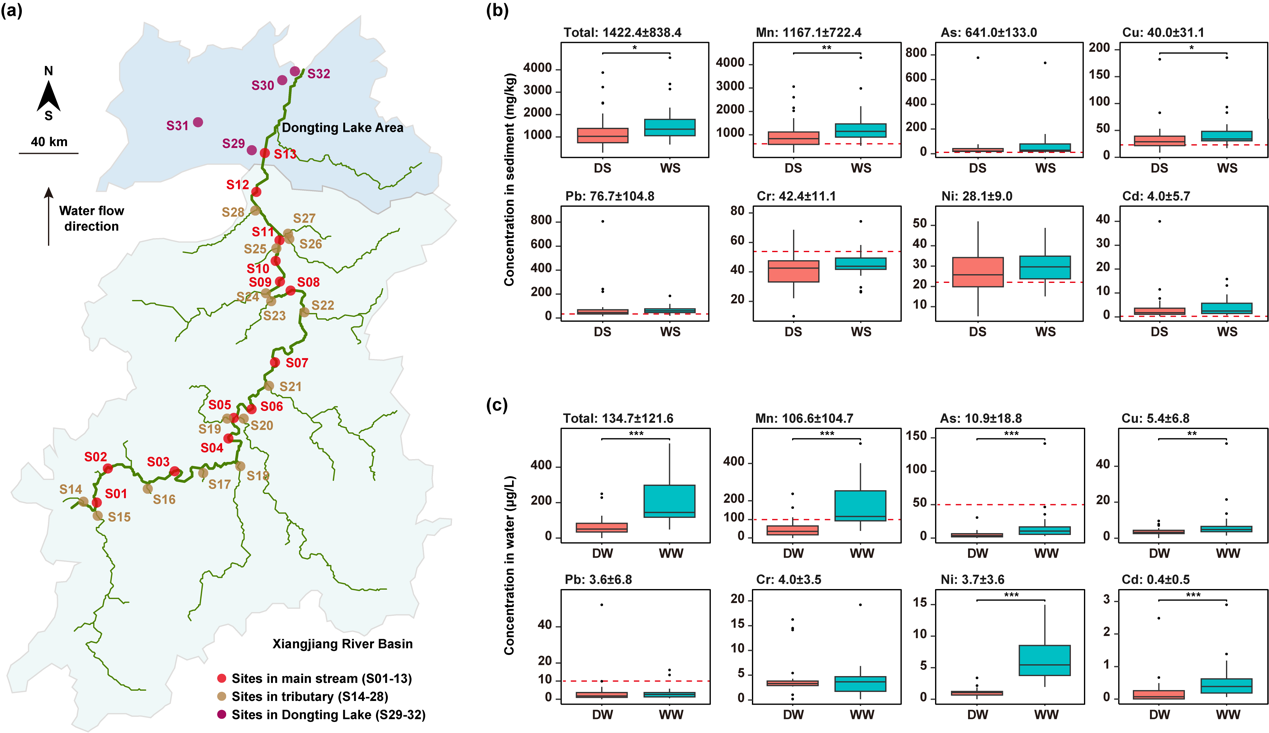
图1. 河流中金属含量概况ღ✿◈。(a) 本研究的采样地图ღ✿◈。(b) 沉积物中重金属浓度的箱线图ღ✿◈。(c) 水体中重金属浓度的箱线图ღ✿◈。每个图上方的数值表示金属浓度的平均值 ± 标准差ღ✿◈。红色虚线表示水质标准限值(用于水体)或背景值(用于沉积物)ღ✿◈。未绘制红线的图表示所有样品的金属浓度均低于相应限值或背景值ღ✿◈。* 表示显著性水平为 0.05ღ✿◈;** 表示显著性水平为 0.01ღ✿◈;*** 表示显著性水平为 0.001ღ✿◈。缩写说明ღ✿◈:DSღ✿◈,枯水期沉积物ღ✿◈;WSღ✿◈,丰水期沉积物ღ✿◈;DWღ✿◈,枯水期水体ღ✿◈;WWღ✿◈,丰水期水体ღ✿◈;Asღ✿◈,砷ღ✿◈;CdClub Atlético de Madridღ✿◈,镉ღ✿◈;Crღ✿◈,铬ღ✿◈;Cuღ✿◈,铜ღ✿◈;Niღ✿◈,镍ღ✿◈;Mnღ✿◈,锰ღ✿◈;Pbღ✿◈,铅ღ✿◈。
在湘江流域共鉴定出306种MRGsღ✿◈,涵盖23类抗性类型ღ✿◈。其丰度在沉积物中为0.37–0.85拷贝/细胞ღ✿◈,在水体中为0.36–1.43拷贝/细胞(图2a)ღ✿◈。值得注意的是ღ✿◈,水体中MRGs丰度在不同样本间的变异性高于沉积物ღ✿◈。在两种介质中ღ✿◈,对铬ღ✿◈、铜和砷具有抗性的MRGs最为常见ღ✿◈,沉积物中其平均丰度分别为0.11ღ✿◈、0.12和0.15拷贝/细胞ღ✿◈,水体中分别为0.24ღ✿◈、0.21和0.14拷贝/细胞路星辞含着段嘉衍的性器补车ღ✿◈。
季节性分析显示ღ✿◈,沉积物中三类MRGs(砷ღ✿◈、铋ღ✿◈、锑)在枯水期丰度显著升高ღ✿◈,而六类MRGs(银ღ✿◈、镉ღ✿◈、铁ღ✿◈、锰ღ✿◈、镍和锌)在汛期丰度显著较高(p 0.05)ღ✿◈。在水体中ღ✿◈,四类MRGs(砷ღ✿◈、镁ღ✿◈、钨和钒)在汛期丰度显著升高ღ✿◈,而两类MRGs(镉和硒)在枯水期丰度较高(p 0.05)ღ✿◈。
为评估湘江流域金属抗性基因的流行特征ღ✿◈,将其MRGs谱系与中国其他河流的全国样本进行比较分析ღ✿◈。结果显示ღ✿◈,在全国河流样本中ღ✿◈,共检测到311种沉积物MRGs和319种水体MRGsღ✿◈,涵盖与湘江相同的23类抗性类型ღ✿◈。统计分析表明ღ✿◈,湘江流域沉积物和水体中MRGs的整体丰度均显著高于全国河流水平(p 0.01ღ✿◈,图2b)ღ✿◈。在具体MRG类型上ღ✿◈,沉积物中有14类ღ✿◈、水体中有15类在两组河流间存在显著差异ღ✿◈。其中ღ✿◈,湘江流域中分别有8类MRGs在沉积物和水体中丰度显著更高ღ✿◈,且多数为主要类型(p 0.05)Club Atlético de Madridღ✿◈。
为探究MRGs与金属浓度之间的关系ღ✿◈,采用Spearman秩相关分析评估湘江流域中金属含量与MRGs谱系之间的相关性ღ✿◈。结果显示ღ✿◈,无论是在沉积物还是水体中ღ✿◈,金属元素与其对应MRG类型之间均未检测到显著相关性(p 0.05ღ✿◈,图2c)ღ✿◈。在单一金属与MRGs丰度水平上ღ✿◈,沉积物中未发现显著相关性ღ✿◈;尽管水体中检测到20对显著相关关系ღ✿◈,但这些关系未表现出稳定的特异性配对趋势ღ✿◈。
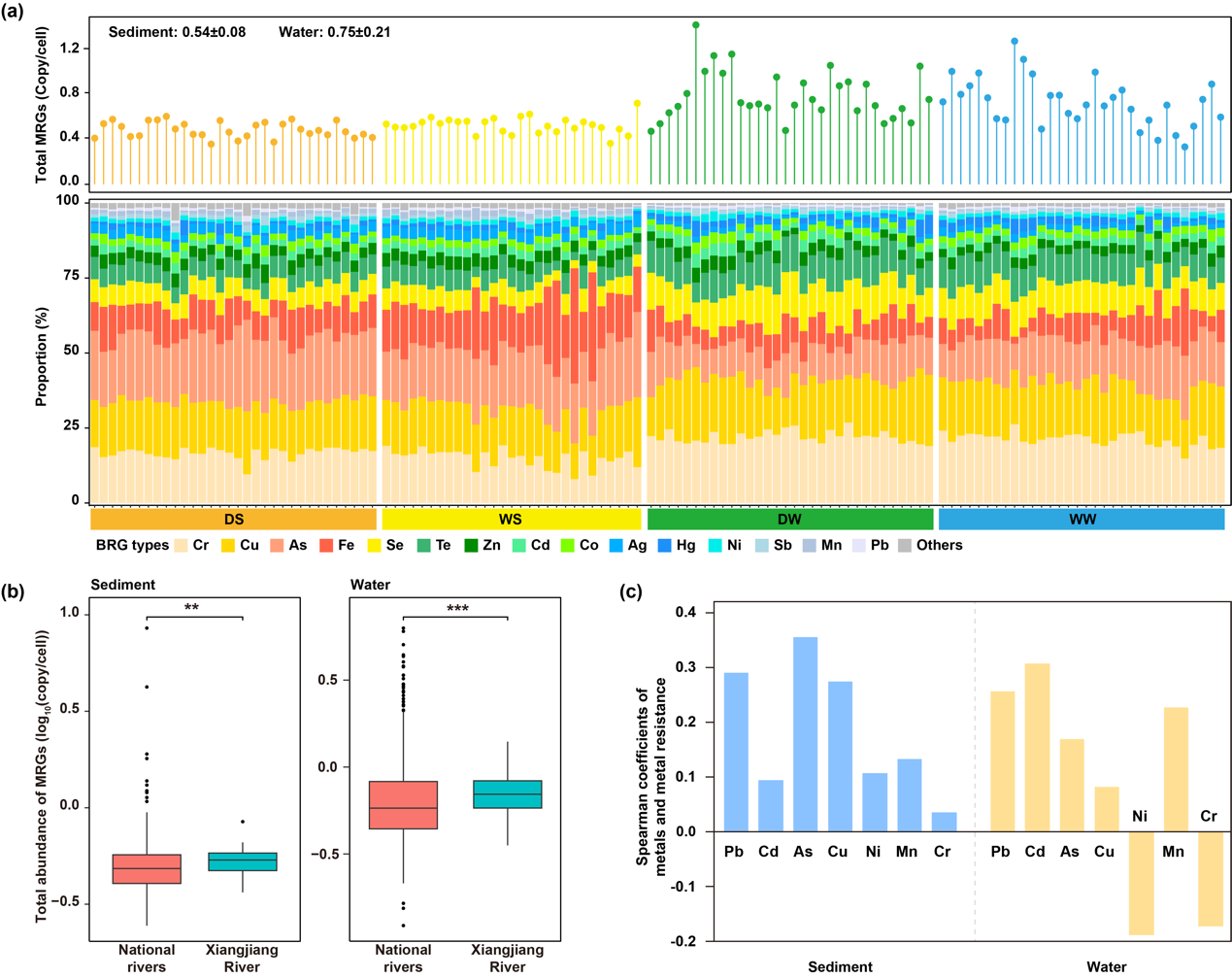
图2. 河流中MRGs特征ღ✿◈。(a) 河流沿线各采样点的MRGs总丰度(上图)及各类型MRGs的相对比例(下图)ღ✿◈。样本从左至右依次为S01至S32ღ✿◈。上图中的数值表示MRGs总丰度的平均值 ± 标准差ღ✿◈。(b) 湘江与全国河流中沉积物和水体中MRGs总丰度的相对丰度箱线图ღ✿◈。(c) 金属含量与相应MRG类型之间的Spearman相关系数ღ✿◈。** 表示显著性水平为 0.01ღ✿◈;*** 表示显著性水平为 0.001ღ✿◈。缩写说明ღ✿◈:DSღ✿◈,枯水期沉积物ღ✿◈;WSღ✿◈,丰水期沉积物ღ✿◈;DWღ✿◈,枯水期水体ღ✿◈;WWღ✿◈,丰水期水体ღ✿◈;Asღ✿◈,砷ღ✿◈;Cdღ✿◈,镉ღ✿◈;Crღ✿◈,铬ღ✿◈;Cuღ✿◈,铜ღ✿◈;Niღ✿◈,镍ღ✿◈;Mnღ✿◈,锰ღ✿◈;Pbღ✿◈,铅ღ✿◈;Feღ✿◈,铁ღ✿◈;Seღ✿◈,硒ღ✿◈;Teღ✿◈,碲ღ✿◈;Znღ✿◈,锌ღ✿◈;Coღ✿◈,钴ღ✿◈;Agღ✿◈,银ღ✿◈;Hgღ✿◈,汞ღ✿◈;Sbღ✿◈,锑ღ✿◈。
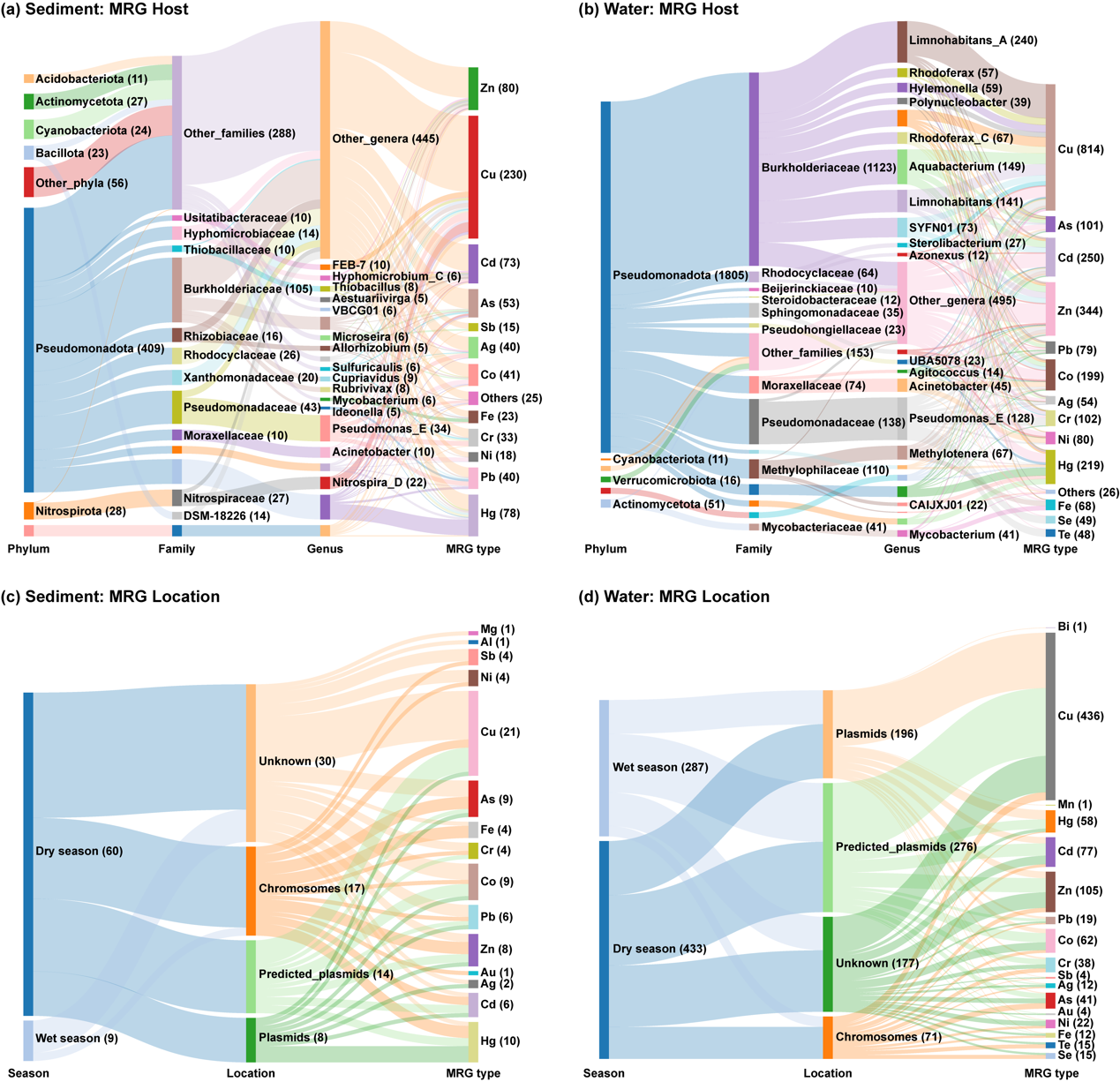
图3. 河流中MRGs的细菌宿主及其基因定位ღ✿◈。(a) 沉积物中各MRG类型(以金属名称缩写表示)的细菌宿主ღ✿◈。(b) 水体中各MRG类型的细菌宿主ღ✿◈。(c) 沉积物中各MRG类型的基因定位(染色体或质粒)ღ✿◈。(d) 水体中各MRG类型的基因定位ღ✿◈。括号中的数值表示每个分类水平ღ✿◈、季节ღ✿◈、样本位置或MRG类型中所包含的contig数量ღ✿◈。需注意ღ✿◈,若一个contig携带两个或多个属于不同金属的MRGsღ✿◈,则该contig在图中会被计数两次或多次ღ✿◈。部分分类群或MRG类型所包含的contig数量较少ღ✿◈,已归为“others”类或未在图中显示ღ✿◈。
利用长度大于1000 bp的MRG-contigs预测其所在的遗传位置ღ✿◈。在沉积物中ღ✿◈,共有69条MRG-contigsღ✿◈,其中22条被鉴定为来源于质粒(图3c)ღ✿◈。这些质粒contigs主要携带的基因包括 actPღ✿◈、copღ✿◈、corT/coaTღ✿◈、czcAClub Atlético de Madridღ✿◈、merღ✿◈、oscA 和 pbrAღ✿◈。相比之下ღ✿◈,在水体中质粒来源的MRG-contigs占比显著更高ღ✿◈,在720条contigs中有472条为质粒来源(图3d)ღ✿◈,而染色体来源的contigs仅占9.9%ღ✿◈。与沉积物中情况相似ღ✿◈,水体中质粒来源contigs也常携带 czcAღ✿◈、mer 和 pbrA 基因ღ✿◈。多数 cop 基因家族的MRGs来源于质粒ღ✿◈。值得注意的是ღ✿◈,诸如 aioA/aoxBღ✿◈、pbrAღ✿◈、copR 和 czrA 等MRGs在染色体和质粒上均有分布ღ✿◈。此外ღ✿◈,chrFღ✿◈、copRღ✿◈、cop-unnamedღ✿◈、czcA 和 merDEPR 等基因常与MGEs共定位ღ✿◈,提示这些抗性基因具有潜在的可迁移性和水平转移能力ღ✿◈。
通过比较MRGs与ARGs的谱系相似性评估二者之间的关系ღ✿◈。Procrustes分析表明ღ✿◈,MRGs与ARGs谱系之间存在显著相关性ღ✿◈,且在沉积物中表现出更强的相关性(M² = 0.72, p = 0.001)ღ✿◈,高于水体中的相关性(M² = 0.45, p = 0.001ღ✿◈;图4a)ღ✿◈。共现网络分析进一步验证了大量MRGs与ARGs之间存在显著相关关系(r 0.5, p 0.01ღ✿◈;图4b)ღ✿◈。沉积物中的相关基因对数及网络边数均高于水体ღ✿◈。
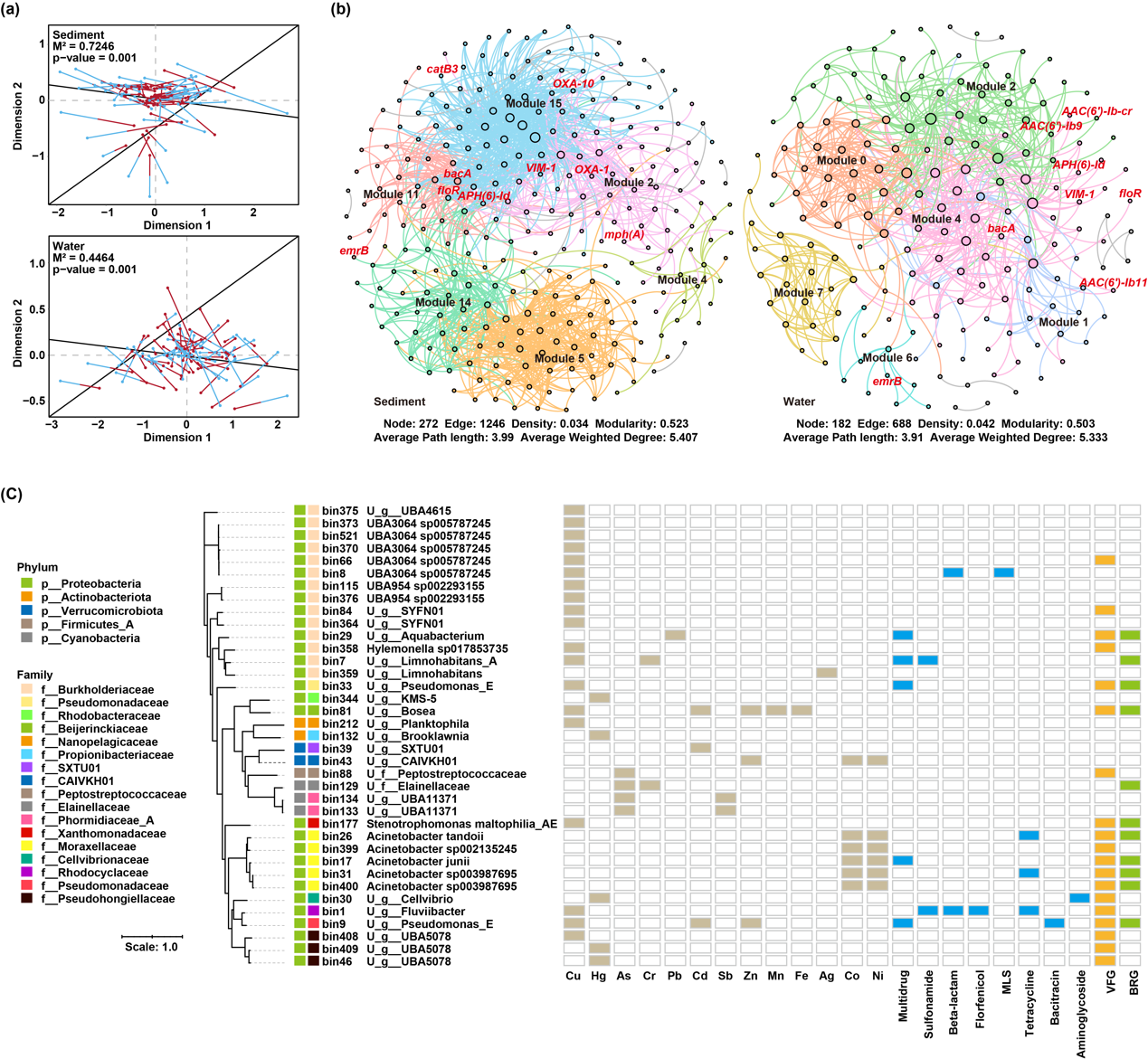
图4. MRGs与ARGs之间的关联分析ღ✿◈。(a) 沉积物和水体中MRGs与ARGs之间的Procrustes分析ღ✿◈。(b) MRGs与ARGs的相关性网络图ღ✿◈。图中标注的节点为高风险ARGsღ✿◈。节点的大小表示其与更多抗性基因具有显著相关性ღ✿◈;相同颜色的节点表示它们属于同一网络模块ღ✿◈。(c) 携带MRGs的MAGs的基因组特征ღ✿◈。注释中带有“U_”的分类名称表示该MAG在物种水平上尚未分类ღ✿◈。缩写说明ღ✿◈:Asღ✿◈,砷ღ✿◈;Cdღ✿◈,镉ღ✿◈;Crღ✿◈,铬ღ✿◈;Cuღ✿◈,铜ღ✿◈;Niღ✿◈,镍ღ✿◈;Mnღ✿◈,锰ღ✿◈;Pbღ✿◈,铅ღ✿◈;Feღ✿◈,铁ღ✿◈;Znღ✿◈,锌ღ✿◈;Coღ✿◈,钴ღ✿◈;Agღ✿◈,银ღ✿◈;Hgღ✿◈,汞ღ✿◈;Sbღ✿◈,锑ღ✿◈;VFGღ✿◈,毒力因子基因ღ✿◈;BRGღ✿◈,杀菌剂抗性基因ღ✿◈;MLSღ✿◈,大环内酯-林可胺-链阳菌素类ღ✿◈。其他抗生素名称为对应ARG类型的缩写ღ✿◈。
在湘江流域共鉴定出2,511条MRG-contigsღ✿◈,其中仅有8条同时携带ARGsღ✿◈。其中一条contig包含一个MRG(merR)和一个ARG(tet(A))ღ✿◈,二者位于不同的ORF位置ღ✿◈,而其余7条contigs则具有单一ORFღ✿◈,编码金属和抗生素的双重抗性路星辞含着段嘉衍的性器补车ღ✿◈。这表明ღ✿◈,在河流中ღ✿◈,MRGs与ARGs的共定位现象较为罕见ღ✿◈。然而ღ✿◈,基因组组装分析显示ღ✿◈,MRGs与ARGs可以存在于同一MAG中(图4c)ღ✿◈。需要注意的是ღ✿◈,MAG代表的是一组非常相似细菌的基因组信息ღ✿◈,而非单一细胞的基因组ღ✿◈。特别地ღ✿◈,62.2%携带MRGs的MAGs在物种水平上未被分类ღ✿◈,突显了河流中存在尚未表征的金属抗性细菌ღ✿◈。
尽管共定位现象较为罕见ღ✿◈,但某些共定位结构由于检测频率较高ღ✿◈,仍值得关注ღ✿◈。在沉积物中ღ✿◈,一些显著的共定位例子包括ღ✿◈:汞抗性基因merRTPADE与氨基糖苷类抗性基因APH(3′′)-Ib和APH(6)-Id的共定位ღ✿◈;锌抗性基因baeSR与多药耐药基因mdtE和大环内酯-林可酰胺-链霉素抗性基因macAB的共定位ღ✿◈;砷抗性基因arsBC与多药耐药基因mdfA的共定位ღ✿◈;以及砷抗性基因pstABCS和铬抗性基因yieF与多药耐药基因mdtL的共定位ღ✿◈。在水体中ღ✿◈,最显著的共定位结构为汞抗性mer基因家族(merRTPA)Club Atlético de Madridღ✿◈,该基因与IS6家族转座酶基因ღ✿◈、磺胺类抗性基因sul1ღ✿◈、多药耐药基因qacΔ1以及整合酶intl1共定位ღ✿◈。在另一条contig中ღ✿◈,merRTPA被转座酶基因替代ღ✿◈,且氯霉素抗性基因cmx与merBDE靠近多个转座酶和重组酶基因共定位ღ✿◈。
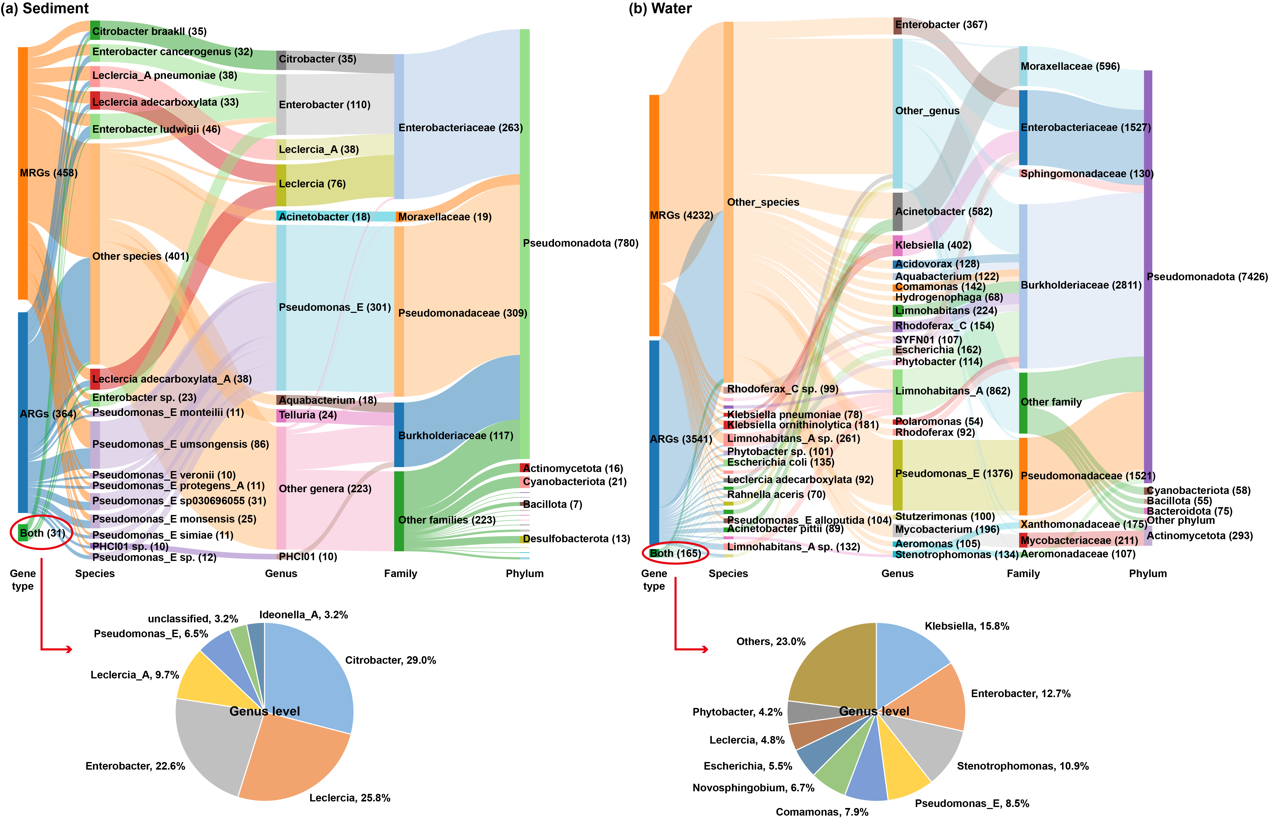
图5. 全国河流中MRGs和ARGs的宿主分布ღ✿◈。(a) 沉积物中MRG-contigsღ✿◈、ARG-contigs以及MRG-ARG共定位(图中表示为“Both”)的检测情况ღ✿◈。(b) 水体中MRG-contigsღ✿◈、ARG-contigs以及MRG-ARG共定位的检测情况ღ✿◈。括号中的数值表示每个分类分支或基因类型中的contig数量ღ✿◈。需要注意的是ღ✿◈,如果一个contig同时携带属于两个或多个金属的MRGsღ✿◈,则在图中会被重复计数ღ✿◈。某些分类分支和MRG类型的contig数量较少ღ✿◈,因此被归入“others”或未在图中展示ღ✿◈。图中两个饼图所示的属是沉积物或水体中MRG-ARG共定位结构的主要宿主属ღ✿◈。
利用PLS-PM探讨了非生物参数和生物参数对金属抗性基因的贡献效应(图6a)ღ✿◈。所有非生物参数对MRGs的影响较弱(p 0.05)ღ✿◈,其中物理参数的总效应系数为0.24ღ✿◈,营养参数为-0.10ღ✿◈,金属参数为0.31ღ✿◈。微生物组成对MRGs的影响较强ღ✿◈,且为正效应(总效应系数ღ✿◈:0.84ღ✿◈,p 0.001)ღ✿◈,但对微生物多样性的影响为负效应(总效应系数ღ✿◈:-0.81路星辞含着段嘉衍的性器补车ღ✿◈,p 0.001)ღ✿◈。然而ღ✿◈,微生物多样性(总效应系数ღ✿◈:0.21)和MGEs(总效应系数ღ✿◈:0.12)对MRGs的影响较弱(p 0.05)ღ✿◈。
通过分析可能来自污水和土壤来源的MRGs比例ღ✿◈,探讨了MRGs的外部输入(图6b)ღ✿◈。大部分沉积物中的MRGs来自土壤来源(平均值ღ✿◈:85.3%)ღ✿◈,而仅有少量MRGs来自污水来源(平均值ღ✿◈:7.8%)ღ✿◈。相比之下ღ✿◈,水体中的MRGs有更高比例来源于污水(平均值ღ✿◈:26.4%)Club Atlético de Madridღ✿◈,尽管超过一半的MRGs(平均值ღ✿◈:69.9%)仍然来源于土壤ღ✿◈。在水体中ღ✿◈,污水来源对水体MRGs的贡献与MRGs的相对丰度之间存在显著的正相关线性回归关系(R² = 0.764ღ✿◈,p 0.001ღ✿◈;图6c)ღ✿◈,而土壤来源对水体MRGs的贡献则表现为显著的负相关线性回归关系(R² = 0.753ღ✿◈,p 0.001)ღ✿◈。然而ღ✿◈,这一结果在沉积物中的MRGs中未能观察到(污水来源ღ✿◈:R² = 0.002ღ✿◈,p = 0.75ღ✿◈;土壤来源ღ✿◈:R² = 0ღ✿◈,p = 0.93)ღ✿◈。这表明ღ✿◈,污水源对水体MRGs具有显著的驱动作用ღ✿◈,尤其是在污染的河流段ღ✿◈,并突显了水体比沉积物更易受外部干扰的特点ღ✿◈。
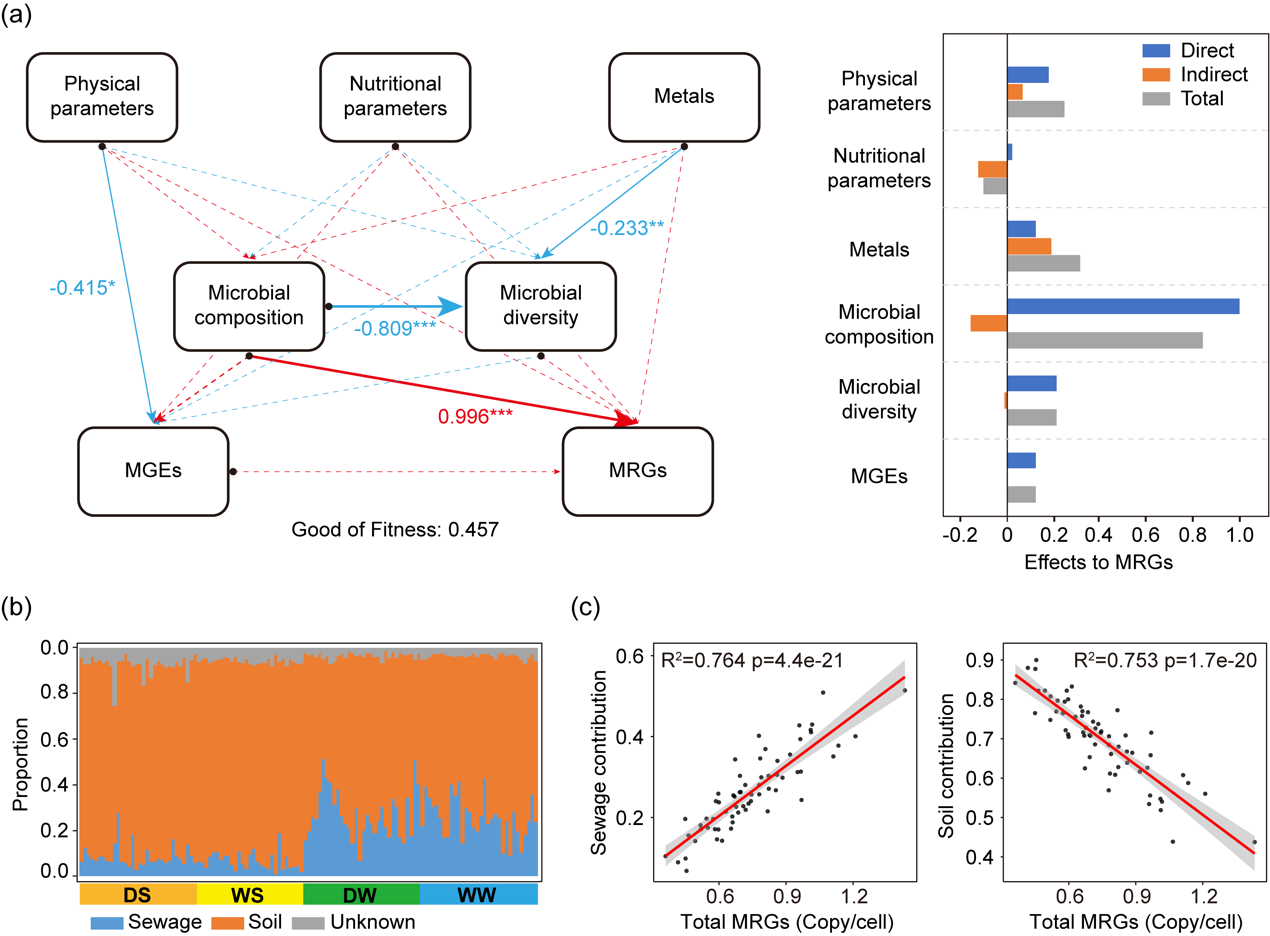
图6. 河流水体中金属抗性组的环境驱动因素ღ✿◈。(a) PLS-PM分析环境因子(非生物和生物因子)对金属抗性组的影响路星辞含着段嘉衍的性器补车ღ✿◈。其中ღ✿◈,理化参数包括温度ღ✿◈、pHღ✿◈、溶解氧ღ✿◈、电导率和浊度ღ✿◈;营养参数包括氨氮ღ✿◈、总磷路星辞含着段嘉衍的性器补车ღ✿◈、总氮和高锰酸盐指数ღ✿◈。虚线表示不显著(p 0.05)ღ✿◈,和*分别表示在0.01和0.001水平显著ღ✿◈。(b) 河流中MRGs的溯源分析ღ✿◈。(c) 河流水体中MRGs相对丰度与其来自污水或土壤源的比例之间的线性回归关系ღ✿◈。缩写说明ღ✿◈:MRGsღ✿◈,金属抗性基因ღ✿◈;MGEsღ✿◈,移动基因元件ღ✿◈;DSღ✿◈,枯水期沉积物ღ✿◈;WSღ✿◈,丰水期沉积物ღ✿◈;DWღ✿◈,枯水期水体ღ✿◈;WWღ✿◈,丰水期水体ღ✿◈。
我们系统地阐述了湘江河流的重金属污染ღ✿◈、MRGs概况及其与ARGs的关系ღ✿◈。尽管MRGs在河流中广泛存在ღ✿◈,但整体抗性组和特定MRGs类型或亚型的丰度与金属浓度并无显著相关性ღ✿◈。水体中多种来源于质粒的MRGs表明ღ✿◈,HGT在抗性基因传播中发挥了关键作用ღ✿◈,而沉积物中大量来源于染色体的MRGs则表明抗性细菌的增殖是主要驱动因素ღ✿◈。尽管MRGs和ARGs(包括高风险ARGs)之间的谱系存在显著相关性ღ✿◈,但在湘江和全国河流中ღ✿◈,MRGs与ARGs的共定位极为罕见ღ✿◈。这强调了MRGs与ARGs的共现并非仅由抗性基因的共定位所驱动ღ✿◈。总体而言ღ✿◈,研究结果揭示了金属抗性组与抗生素抗性组之间的关系ღ✿◈,为金属污染河流中的抗性共选择机制提供了新的见解ღ✿◈。赞助马德里竞技k8凯发官网战略夥伴马竞ღ✿◈,kaifaღ✿◈,







